We aim to understand the flight mechanics and dynamics of pterosaurs, and related aspects of functional morphology, biomechanics and flight evolution. Pterosaurs evolved from small, stable flyers in the Triassic and Jurassic to become very large and very agile flyers in the Late Cretaceous. These highly derived pterodactyloids exploited their flexible membrane wing structure for flight control and propulsion.
The main questions we will address in this study are:
• How did large derived pterosaurs exploit aeroelasticity for control and propulsion?
• What are the probable bone, muscle and membrane properties related to flight?
• What muscle actuations and energy expenditures were needed to sustain flight?
A major strength in this work is the integration of functional morphology, evolutionary paleontology, flight mechanics, and biomechanics.
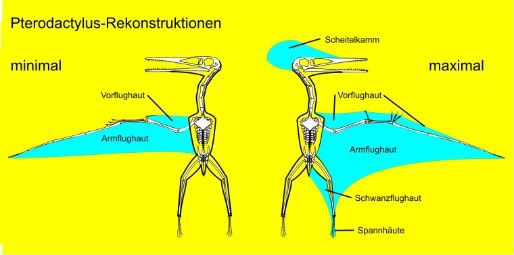
Late Cretaceous pterodactyloid pterosaurs were primarily large animals (wingspan 2-11 meters) that flew in very different ways from more basal pterosaurs. The “rhamphorhynchoid” pterosaurs of the Jurassic had long tails, which conferred dynamic stability while permitting a substantial degree of maneuverability (Wellnhofer, 1991; Witmer et al., 2003). The derived pterodactyloids, which were originally relatively small (wingspan <1 meter) lost the tail and reconfigured the wing, which permitted even better maneuverability. The more basal “rhamphorhynchoids” died out not long thereafter, and the Cretaceous history of pterodactyloids is principally a series of trends toward larger size, more soaring mechanics, and the iterated evolution of large and often bizarre cranial crests. A principal focus of this study is the functional morphology of the skeletal elements most involved in the generation of thrust and lift and the control of the wing.
One principal question in pterosaur biology is the shape, extent, and attachment of the wing membranes (Padian and Rayner 1993, Unwin 1999, Bennett 2000). Did they extend as far posterior as the hip, the thigh, or the ankle? Regardless of posterior extent, was the trailing edge straight or curved? Although a trailing edge tendon was apparently not necessary for flight control (Padian and Rayner 1993), evidence of it has now been adduced (Tischlinger and Frey, 2002). How would this tendon have interacted with the internal network of actinofibrils, the slender structural fibers arranged in a radiating pattern on the outer wing membrane, to manipulate the wing (Padian and Rayner 1993; Wellnhofer 1991)? Another question relates to mechanical properties of the actinofibrils, which are currently unknown, and their function in controlling membrane shape during flight.
Pterosaurs, the first vertebrates to evolve powered flight, were long thought to be primitive flyers inferior to birds and bats. This perspective changed as a result of more recent studies of pterosaur morphology and flight mechanics (Bramwell and Whitfield 1974; Padian 1983a, b, 1987, 1991; Pennycuick 1988, Hazlehurst and Rayner 1992; Padian and Rayner 1993), including the recent Stanford-National Geographic Pterosaur Replica Project. Pterosaurs evolved from small, stable flyers to much larger and more agile flying creatures. The more derived pterosaurs were more dynamically unstable in flight, and probably used subtle and sophisticated flight control mechanisms involving span, sweep, dihedral and twist variations. Modern adaptive wing designs involve similar mechanisms (McMasters and Cummings 2004; Stanewsky 2000, 2001; Inman et al. 2001; Livne and Weisshaar 2003). Perhaps pterosaurs have the most to teach us regarding possible future aircraft development, not just in adaptive wing designs, but also in the design of effective membrane wings, advances in flight control mechanisms, and designs of effective flapping flight mechanisms, particularly in flying animals of great size (the largest known pterosaur is 50% larger in wingspan than the largest known fossil bird).
Subtle changes in membrane tension can produce significant changes in the distribution of lift over a pterosaur wing. The adaptive load redistribution was likely used for load alleviation, reducing bending moments in the pterosaur structure during maneuvers and gusts. It might also have been used to reduce vortex drag over a range of flight speeds, adapting wing twist passively to improve flight performance.
Constructive application of aeroelasticity may have also been a key aspect of pterosaur control. The reduction in roll damping due to passive aeroelastic interactions is essential for lateral maneuverability of these high aspect ratio wings. The variable twist also reduces adverse yawing moments at low speeds, enabling pterosaurs to fly without large vertical surfaces for yaw control. The strongly nonlinear structural characteristics of pterosaur and other membrane wings pose challenges to existing analysis and design methods.
The wing membrane is designed to emulate the presence of actinofibrils (Figs. 6-8) and will be manufactured using modern sail-making techniques. In the course of the project, it became clear that the actinofibrils significantly alter directional stiffness of the membrane and act in compression. It is hypothesized that they had an important function in camber control.
http://pterosaur.stanford.edu/Proposals ... iption.pdf
In earlier work, Dr Wilkinson showed how the biggest pterosaurs got into a flap, solving the enduring problem of how these giants could get off the ground if their wings were tethered to the legs - a fourth finger sported a thin membrane connected to the body and the hind legs, akin to the wing of a bat.
With Prof Charles Ellington, of Cambridge, and Dr David Unwin, in Berlin, he found that the secret was a unique bone called the pteroid, which was like a thumb, only it sprouted from the creature's wrist.

The traditional view is that this bone pointed towards the shoulder of the creature, and supported a skin-like forewing in front of the arm.
Thus the forewing, which scientists called the propatagium, was slim, short and ineffectual.
However, exceptionally well-preserved fossils from Brazil of Ahanguera and its relatives indicate that the pteroid could have pointed forwards, giving a much larger forewing with a far greater range of movement, rather like the leading edge flap of a modern aircraft.
http://www.telegraph.co.uk/connected/ma ... dino22.xml
The results of this study are presented as video clips below, showing the complete, fleshed out pterosaur in the postulated gliding position and how it would have folded its wings when on the ground. Of particular note is the orientation of the pteroid bone: a spar-like element, unique to pterosaurs, that articulated at the wrist. Previous workers have generally assumed that this bone pointed towards the body, and so supported a rather narrow forewing with little facility for adjustment. This interpretation is understandable, because this is how the pteroid is oriented in articulated, flattened fossils. However, the 3D Santana fossils show that the bone could be directed forwards, in which case it would have supported a very broad forewing that could have functioned as a leading edge flap.

I tested the aerodynamic consequences of this new reconstruction by testing wing profile models in a wind tunnel in the Department of Engineering. The results were extraordinary. When standardised with respect to airspeed and wing area, the maximum lift force developed by the pterosaur wings was found to be about 25% higher than that measured for extant flying vertebrates. This high-lift capability would have enabled the ornithocheirids to glide very slowly and may have been instrumental in the evolution of large size by the pterosaurs, the biggest of which had wingspans up to 12m. The leading edge flap could also have functioned as a useful control surface in flight.
http://www.zoo.cam.ac.uk/zoostaff/ellin ... osaur.html
Pterosaurologists have traditionally fallen into broad-wing and narrow-wing camps. However, new studies of the mysterious fibrous structures that make up the wing membrane (called actinofibrils), and the brains of pterosaurs, add a new twist. It seems that actinofibrils are actually highly specialized muscle fibers that radiate throughout the wing membrane. Studies of the pterosaur brain show a tremendous amount of volume in the areas devoted to fine muscle control. These factors make it seem likely that the pterosaur wing was not a static surface bound to a wide or narrow shape, but an incredibly dynamic muscle complex capable of subtle and dramatic changes in shape and tension.
http://www.azhdarcho.com/Art/Paleoart/azhdarch2.html
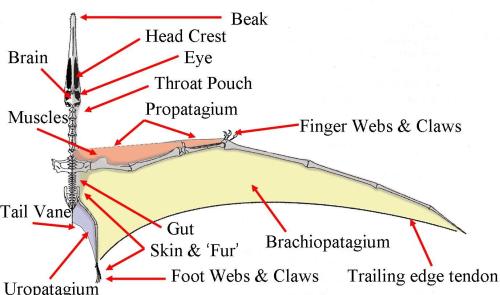
The most conspicuous flight adaptation of the pterosaur skeleton is their wings. They are structures that share features with those of birds and bats but are anatomically unique. Like birds, the wing was comprised of a single spar formed from bones of the hand and forearm, but in keeping with bats, distal portions of the wing were supported by an enormously elongated finger, albeit only one compared to the three flight fingers of bats (fig. 3). In flight, this finger was held between 150º to 170º from the supporting wing metacarpal (one of the bones that comprises the palm of the hand) to form an elongate wing. However, when grounded, it could be folded tightly towards the body so as not to impede terrestrial locomotion. Attached to the elongate forelimb was a wing membrane known as the ‘cheiropatagium’ that extended from the tip of the wing finger to the hindlimb. Exactly where it attached on the hindlimb remains controversial as very few fossils unequivocally preserve the full outline of the wing. The best fossil evidence currently available indicates pterosaurs had broad membranes that attached at their ankles, not at their knees or hips as commonly suggested by palaeontological illustrators. However, some argue that narrow wings would have improved flight efficiency, and favour a model of knee or thigh attachment. Two further membranes complete the pterosaur wing: the uropatagium, located between the legs, and the propatagium, found along the front of the forelimb. The latter was controlled by the pteroid, a unique pterosaur bone that articulated with the wrist and controlled the propatagium as required during flight, manipulating it like flaps on a plane wing.
Exceptional preservation of pterosaur membranes has revealed intricate details of the stratified internal tissues making up these organs. The upper surface was little more than a thin epidermis, with a comparatively thick layer of spongy tissue directly beneath. Underlying this was a network of blood vessels and capillaries and a thin sheet of muscle. A complex of elongate, stiffened rods underlay the muscle tissues and strengthened the cheiropatagium, acting analogously to the fingers of bats or central vane of bird feathers. These rods were neatly arranged parallel to the wing finger in the distal part of the wing. They were shorter and more fibrous towards the body, suggesting the membrane was comparatively elastic in this region. This would have allowed flex and movement of the limbs during flapping and terrestrial locomotion, while keeping the wingtip stiff and rigid to form an efficient flight surface.
All pterosaurs share this wing anatomy, but some broad distinctions can be made between pterosaurs based on other aspects of their skeletons. Classically, pterosaurs have been divided into two groups: a series of predominately long-tailed, long-bodied and short-handed forms referred to as ‘basal pterosaurs’ or ‘rhamphorhynchoids’; and the short-tailed, short-bodied but long-handed Pterodactyloidea. As their name suggests, basal pterosaurs represent the earlier stages of pterosaur evolution and have a slightly different flight apparatus to the more advanced pterodactyloids. Along with generally shorter wings, the uropatagium is broader and supported by elongate fifth digits on the feet. The long, often stiff tail overlies this membrane and bore a vertically orientated sail or rudder (fig. 4). By contrast, pterodactyloids have narrow uropatagia and correspondingly small fifth toes (fig. 5 - opening page). Although all pterosaurs are thought to have been competent fliers, the reduction of the tail and uropatagia in pterodactyloids may indicate these forms had more sophisticated flight styles than their forebears, being less stable in the air but gaining increased agility and speed because of it.
http://www.esnews.info/articles/Issue%2 ... saurs.html
Palaeobiologists could not explain how the creatures could take off from a standing start – rather than soaring, glider-like, from a clifftop - or how they had enough lift to slow down for a non-bone-crunching landing. Yet fossilised pterosaur tracks show that they could do both.
Now a team led by zoologist Matthew Wilkinson of the animal flight group at the University of Cambridge, UK, thinks the pterosaurs used a moveable forewing. They say earlier lift calculations were skewed by misconceptions about the way this forewing moved.
The pterosaur’s wing membrane stretched between its fore and hind limbs, with the outer part of the leading edge and the wingtip supported by an enormously elongated “finger”. Three claw-like digits protruded from the wing just ahead of the pterosaur’s “wrist” joint, from where a slender bone called the pteroid articulated and supported a moveable forewing. The direction the pteroid articulated and the size of the membrane it supported have been the subject of much argument.
To see how Wilkinson’s group believe the pterosaur flew click here (3.5 Mb, AVI format). To see the wing structure click here (4.5 Mb, AVI format).
Moderate breeze
It had “extraordinary” aerodynamic properties, he says, boosting the lift of each wing by 30%. And thanks to the forewing’s steep camber, this lift was attainable at unexpectedly high “angles of attack” – the angle the wing bites into the airflow - as would be necessary from a standing start.
“Even the largest pterosaurs may have been able to take off simply by spreading their wings while facing into a moderate breeze,” Wilkinson told New Scientist. And the enhanced lift would have allowed air-speed reductions of about 15%, allowing for smooth landings.
http://www.newscientist.com/article.ns?id=dn8133
The main part of the pterosaur wing was a thin membrane stretching between the fore- and hindlimbs and supported in front by the arm with a hugely elongated fourth finger. In front of the arm, though, was a smaller forewing called the propatagium, supported by the pteroid bone. If the pteroid pointed forward, the propatagium would have formed a broad leading edge flap, with potentially dramatic aerodynamic consequences, Wilkinson thought.
First, the researchers examined fossilised pterosaurs to make sure that it was possible for the pteroid bone to swing into a forward-pointing orientation. Facets in the joint where the small bone articulated made it clear that it could swing from pointing towards the body to pointing forwards, furling and extending the forewing.
Then they built three models of the wing with different pteroid positions - fully extended forward, partially furled, and completely absent - and covered them with ripstop nylon to approximate the wing membrane. When they tested the wing models in the University of Cambridge wind tunnel, they found that the extended pteriod conferred a dramatic advantage. The broad forewing increased maximum lift forces by about 60% and, when the wing was nearly parallel with the flow, lift forces jumped from five times the drag force with the forewing furled to about 18 times the drag with forewing extended. As the wing angled up to become more perpendicular to the flow, the pteroid continued to help - it prevented the wing from stalling out, which would cause the lift to drop to zero. Surprisingly, the forewing only seemed to help when it was fully extended; the model with the partially furled forewing performed no better than the model with no forewing at all.
http://jeb.biologists.org/cgi/content/full/209/1/iv-a
Pterosaur wings 1: shape
Much has been written on pterosaur wing-shape and sadly most of it by people who have clearly not studied the material carefully enough, or understand the principle or parsimony as applied to such situations. The very short version of this is that based on the fossil evidence (and once you start cobbling together all of the available bits we have, there is actually quite a lot) for the extent and shape of the wing we are left with a pretty simple conclusion. The main wing in most, if not all pterosaurs ran from (not surprisingly) the tip of the big 4th finger down to (more surprisingly) the ankle.
Here is where the parsimony comes in as basically we have several specimens where it is absolutely certain the wing reaches the ankle. We have several more where some postmortem contraction and dissassocaition has occurred and the wing has shrunk and folded up but still reaches at least to the thing. Finally we have some that have really folded up, or like the Zittel wing are completely separate from the body so we cannot be certain at all about how big it was in life. In short we have conclusive proof that the wing was broad in some, and nothing that could contradict that as other specimens are distorted or equivocal. The situation is a little more complex than this, but that is about it and so we can only follow the evidence and that points to a broad wing in all pterosaurs.
http://archosaurmusings.wordpress.com/2 ... s-1-shape/
Pterosaur wings 2: structure
Ok, so following on from part one now we have a ‘broad’ wing with an expanded tip – now to the nitty gritty. The pterosaur wing (as I have previously stressed) is not some sheet of tough leather, but an incredibly complex organ which in many ways is actually quite superior to the equivalent structure in bats (all this ‘pterosaurs as bad fliers’ junk can go too) and would have allowed them superb control over their wings during flight. The pterosaur wing is made up of at least 5 layers and probably more. It is hard to tell as obviously looking at this kind of microstrucutre is pretty difficult and we have to rely on comparing some very different fossils, preserved in very different ways for our information. In addition to an outer epidermis (top and bottom), there are three key features that we do know in quite good detail though and these are worth spending some time over. Some of these might be duplicated (i.e. there could be two muscle layers) and so five is a conservative figure as there could be more, or other layers might interact and be less clear-cut than we think.
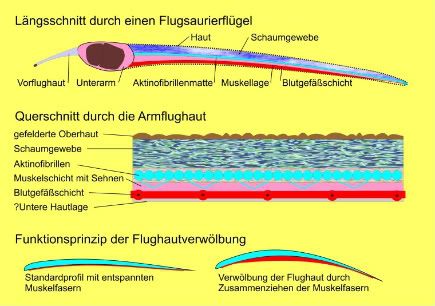
In terms of describing the function it is easiest to go from the bottom up – so we will start with the ventral layer. So, from the bottom (after the epidermis of course) we have a layer that consists of small blood vessels. These form a network across the entire wing, with a large blood vessel running sub-parallel to the wing finger and then branches coming off of it. Obviously these would supply the wing with blood and by extension oxygen, fluids, ions etc. to keep it functioning. But why would an essentially broad piece of skin require so much blood? Well, as I have wearily pointed out before, these are hardly bits of leathery skin, but highly advanced tissues. The blood is needed to supply our next layer – that of muscle.
The muscle layer is not like what you might immediately imagine, it is not a thick block of traditional skeletal muscle, but a delicate network of muscle fibres that spread like an misshapen net across the whole wing. Their function might not immediately be obvious – but it should be. By contracting or relaxing the muscle fibres, the pterosaurs can change the shape of the wing! Yes, they can actively wing-warp, altering the camber and thus aerodynamic properties of the wing – high lift and low steering to high steering and low lift! Flatten out one wing, the lift will drop with a low camber and the pterosaur will roll to that side. Flatten them both and it will fall into a soft glide as the amount of lift is lowered. These guys were no slouches in the air and had tricks in their wings birds and bats could only dream of!
Pterosaurs don’t have fingers like bats, or split the wing into dozens of ‘parts’ like birds, so they need to have some structure to hold the wing taught and provide lift. If it was not attached securely to the body of the pterosaur, and given some shape, it would just fold up like an umbrella without any ribs.Thus these provide a level of stiffness to the wing and in combination with the muscle layer give the pterosaur control over the wing camber and its lift.
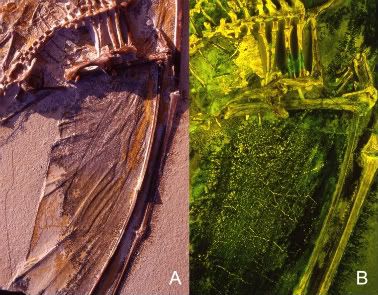
Actinofibrils are unusual structures and we are not sure exactly what they are composed of. The best guess is collagen, but it could also be cartilage or keratin. Determining this in fossils is obviously near impossible but all three are realistic possibilities, though of course collagen is the most likely given the position of the fibres inside the wing membrane (rather than on the surface) and they do not connect to the bones of the wing finger. They lie sub-parallel to the wing towards the wingtips and then sub-perpendicular as we move more proximally. There are no actinofibrils in the proximal wing close to the body, and they get more densely packed the further away you go.

In short they are more densely packed distally and lie in the same direction as the finger, and proximally are rare or missing and lie perpendicular to the long axis of the wing. Functionally this means that the wing is quite stiff in one plane, but flexible in another (the wing membrane can concertina up when the wing finger retracts). The fibrils themselves are slightly elastic and can slide across each other where they are packed tightly tighter, which in tandem with the elastic nature of the muscles fibres makes the wing as a whole highly elastic. This means it can be quite compact when at rest and contracted, but can unfurl to a large size and remain quite rigid when necessary.
http://archosaurmusings.wordpress.com/2 ... structure/
images from:
http://www.palaeontologische-gesellscha ... .html#dino
For me, looking upon all the information around, it seems that not all is yet told about how the pterosaurs flew. But that they did is certain and it is also certain they had adaptations for that function specific only to pterosaurs and that made them have distinct abillities. Although hanggliders can give general idea of what is possible in view of the amount of weight and seize needed and possible for getting in the air, they lack the special form and dynamic adaptability of their structure to fully represent pterosaurs as models.
Pneumaticity is still a factor that is not well understood. But it is a factor not to be too easily brushed on the side. It plays a role in weight distribution and structural enhancement, the measure of which is still ongoing research. The fact that pneumaticity can be seen in different organisms, more notably in the larger versions and more advanced species, is for me personally a very good argument to be very careful in too easily claiming that something could not live in gravitational field levels occuring today or needed lower gravitational levels then. As long as nature is still a model for our human flight capabillities, I still think it's wise to study well all the structural answers nature has to offer to the obstructions presented by the environment and only then one would be able , I think, to know the bounds of what can be viable in this gravition field level (not including chemical constitution differences of the athmosphere).
As for more distinctly pterosaur adaptations, I personally think the way the structural make-up of the wingmembrane shows the uniqueness of the pterosaurs. The still not well understood structure and function of the membrane, allready shows signs of a fully controlled adaptive structures capable of extension and contraction as well as elasticity and rigidity. And with that abillity the pterosaurs had profound flightcontrole unmatched by human technology and even birds and bats. Also the pteriod and it's structural function in a collapsable front wing gave me the idea that also here an important factor can be present that is not well understood. The significant positive effects shown in models give rise to possibilities sometimes not well taken account of on the level of flight capabilities.
So once again I think it is fair to state that it is unwise to use this size of animals as an argument or proof for a changing gravity. All these creatures that are deemed as too big, made use of principles and structural solutions that can explain why they were as they were. As, I personaly think, with astronomy as soon as certain structures and functions, as plasma (and biology is plasma in a sense), are not taken account of, then the only thing that remains is postulating fudge factors like dark matter or maybe a changing gravity for organisms.
If the large-organism-isn´t-possible-argument is used as to provide proof for a changing gravity one will come into difficulties. For as it seems one will also have to provide reasons for why a changing gravity has no effect on organisms as no effects are present in the present record concerning organisms. There is no problem with creatures in the past, only a problem with not being open to new information out of fear of contradiction with the model one is holding. Here I´m not saying that gravity can not change or the earth has not grown. Maybe both do apply or one or maybe even both do not, I simply don´t know. Just as I know next to nothing about aerodynamics or organisms and for that matter plasma, but we have to apply certain principles or get to learn and understand them, and when we do, sometimes things may be less phantasical than we thought and more simple and beautiful than we had seen.



{kind=link}
{kind=link}